





Saimnieks LV / 2020.gads (Septembris.)
Drozofilas ir augļu mušiņas, kas bieži ir redzamas vasarā, kad ir daudz pārgatavojušos un jau pūstošu augļu un ogu. Augļu mušām ir vairākas sugas, viena no tām – punktspārnu augļmuša – ir kļuvusi par bīstamu kaitēkli vairākās Eiropas valstīs. Punktspārnu augļmuša Drosophila suzuki ir potenciāli jauns kaitēklis arī Latvijas augļu dārzos.
Punktspārnu augļmušas raksturojums un ierobežošanas metodes
Kaitēkļa latīniskais nosaukums ir Drosophila suzuki, angliskais – spotted wing drosophila. Tēviņi ir 2.6−2.8 mm gari, ar melnu punktveida plankumu tuvu katra spārna galam. Mātītes ir 3.2−3.4 mm garas un bez šāda punktveida plankuma (EPPO, 2011b). Atšķirībā no citām drozofilām šī augļmuša, izmantojot zobainu dējekli, dēj olas zem mizas ogās un augļos, kas vēl tikai nogatavojas. Vienā dējekļa dūriena vietā mātīte iedēj vienu līdz trīs olas, bet savas dzīves laikā tā var izdēt līdz 300 olām. Tā kā vienu augli olu dēšanai var izmantot vairākas mātītes, tad tajā var attīstīties līdz pat 60–70 pēcnācējiem. Izšķīlušies kāpuri (līdz 3.5 mm gari) un vēlāk arī kūniņas attīstās augļu un ogu iekšpusē, izraisot būtiskus bojājumus, kam var sekot puve. Kūniņas var attīstīties arī augļu un ogu ārpusē. Dzīves cikls ir īss – viena līdz divas nedēļas, atkarībā no klimatiskajiem apstākļiem.
Japānā šim kaitēklim gadā var būt līdz pat 13 paaudzēm, Amerikas Savienotajās Valstīs (Kalifornijā) – 3−10 paaudzes. Tādējādi ir būtiski ražas zudumi. Līdz pagājušā gadsimta 30. gadiem šis kaitēklis bija sastopams tikai Japānā, bet pēdējās desmitgadēs invazīvi izplatījies visos kontinentos, izņemot Antarktīdu. Tas ir konstatēts Sibīrijas dienvidos Krievijā (2003), Spānijā (2010), Itālijā (2010), Francijā (2010) un Slovēnijā (2011). Ir aprēķināts, ka Kalifornijā, ASV, laika periodā no 2008. gada līdz 2014. gadam punktspārnu augļmušas radītie kaitējumi ir sagādājuši 39 milj. USD lielus zaudējumus, sabojājot 20−100 % ražas (Buchman et al 2018; EPPO, 2011a, EPPO, 2011b). Pēdējos gados šis kaitēklis ir konstatēts jau vairākās mums ģeogrāfiski tuvākās valstīs: Polijā (2014), Zviedrijā (2014), Ukrainā, 2014, Somijā (2019) (https://gd.eppo.int/taxon/DROSSU/distribution). Tam patīk klimats ar augstu mitrumu un mērenām temperatūrām. Aukstas ziemas neietekmē kaitēkļa izdzīvošanu, jo tas ir ieviesies gan Ķīnas ziemeļos, gan Hokaido dienvidu daļā (EPPO, 2011b).
Visbiežāk šī augļmuša bojā avenes, kazenes, zemenes, saldos ķiršus, persikus un aprikozes. Retāk tiek bojāti āboli, plūmes un vīnogas. Tas var invadēt arī savvaļas augus, piemēram, mellenes. Šie augļi un ogas, kā arī to tirdzniecība tiek uzskatīti par galvenajiem kaitēkļa pārneses ceļiem no vienas valsts uz citu, jo novākšanas laikā bojājumi var būt vēl neizteikti un nepamanāmi. Kaitēkļa attīstība turpinās transporta laikā. Vēlāk, jau citā valstī, bojātie augļi un ogas var nonākt kompostā, no kurienes pieaugušie īpatņi var invadēt vietējos dārzus. Tirdzniecība ar augu stādāmo materiālu tiek uzskatīta par zema riska izplatības ceļu (EPPO, 2011a, EPPO, 2011b).
Pašlaik pieejamās D. suzukii ierobežošanas metodes balstās uz plaša spektra insekticīdiem, no kuriem neviens Latvijā nav reģistrēts lietošanai pret augļmušām (VAAD, 2020). Valstīs, kur šādi līdzekļi, piemēram, malations, ir reģistrēti, novērota variabla efektivitāte, apgrūtinoša lietošana augļu nogatavošanās laikā, kā arī iespējama D. suzukii rezistences rašanās (Buchman et al 2018). Spānijā ir konstatēti punktspārnu augļmušas dabiskie ienaidnieki – parazitoīdi, spožlapsenes Pachycrepoideus vindemmiae un tumšlapsenes Trichopria cf. drosophilae, kas savā attīstībā izmanto augļmušas kūniņas un būtiski samazina pieaugušo īpatņu izveidošanos bojātajos augļos un ogās. Pētījumā tika konstatēts, ka dabiski ar šiem parazitoīdiem bija invadētas 4−10 % drozofilu kūniņu, bet laboratorijas apstākļos līdz pat 83 %. Atklāti arī plēsīgi kukaiņi no koku laupītājblakšu dzimtas (Orius laevigatus) un spīļastes Labidura riparia, kas barojas ar augļmušas olām vai kāpuriem. Laboratorijas apstākļos šie plēsīgie kukaiņi nogalināja līdz pat 76 % drozofilu olu un kāpuru (Gabarra et al., 2015). Tomēr bioloģiskās kontroles preparāti vēl netiek plaši izmantoti (Buchman et al 2018).
Punktspārnu augļmušas ierobežošana ar gēnu rediģēšanas palīdzību
Viens no variantiem nākotnē šī kaitēkļa populācijas ierobežošanai ir izmantot ģenētiski modificētus organismus, kas izveidoti, balstoties uz gēnu dziņa (gene drive) sistēmām, kuru iedzimtība nenotiek pēc Mendeļa likumiem – gēnu dziņa organismu jeb gēnu sastopamība katrā paaudzē pieaug pat tad, ja nav izdzīvošanas priekšrocību. Viena no vislabāk izstrādātajām sistēmām D. suzukii ierobežošanai ir Medea sistēma (Medea – Maternal Effect Dominant Embryonic Arrest). Medea sistēma balstās uz toksīna – pretlīdzekļa kombinācijas ekspresiju (Buchman et al 2018). Medea sistēma ir dabiski sastopama dabā. Tā ir atklāta miltu vabolei Tribolium castaneum, kur tā regulē populācijas blīvumu, kā arī pelēm Mus musculus, kur tā izraisa smagu kombinētu anēmiju un trombocitopēniju (citēts pēc. COGEM, 2018).
Punktspārnu augļmušai Medea sistēmā microRNS (miRNS) toksīns tiek ekspresēts ooģenēzē sievišķajiem īpatņiem, kuriem ir Medea sistēma, savukārt pretlīdzeklis ekspresējas agrīnās embrioģenēzes stadijās pēcnācējiem, kuriem ir šī sistēma (1. attēls). Toksīnu kodējošais gēns tiek pārmantots visiem pēcnācējiem no mātītes, kurai ir Medea sistēma, kas izpaužas kā miRNS vadīta svarīga ebrioniskā gēna apspiešana, kas izraisa normālas embrioģenēzes attīstības pārtraukšanu. Pēcnācēji, kas pārmanto Medea sistēmu, saņem pretlīdzekli, ko veido šī svarīgā embrioniskā gēna kopija, kas ir rezistenta pret miRNS, atļaujot notikt normālai attīstībai. Pēcnācēji, kuriem nav Medea sistēma, nevar normāli attīstīties un iet bojā. Balstoties uz šādu iedzimtību, Medea sistēma konkrētajā kaitēkļu populācijā ātri izplata pati sevi un ikvienu tajā ievietoto gēnu (Cargo). Šāds ievietotais gēns varētu būt gēns, kas izraisa uzņēmību pret kādu konkrētu ķīmisku vielu un ko aktivē konkrēti vides apstākļi, piemēram, temperatūra vai diapauze (Buchman et al 2018).
Viena no šādām sistēmām tika radīta jau 2007. gadā citai drozofilas sugai – Drosophila melanogaster (Chen et al., 2007). Tika izmantots gēns myd88, kuru pārmanto pa mātes līniju. Šis gēns ir nepieciešams agrās embrija attīstības stadijās, lai nodrošinātu dorzoventrālu attīstību. Ar divu miRNS palīdzību, kas ekspresējas mejozes laikā, šī gēna darbība tiek apturēta. Pretlīdzeklis ir miRNS–nejutīgs myd88 transgēns, kas aktivējas zigotas stadijā (Chen et al., 2007). 2018. gadā šāda sistēma tika radīta punktspārnu augļmušai, izmantojot četras miRNS molekulas, kas iedarbojas uz myd88 gēnu. Iedzimtība nākamajās paaudzēs bija 87−100 %. Tiek uzskatīts, ka Medea sistēmas izmantošanas gadījumā dabā būs nepieciešams izlaist daudz mazāk laboratorijā pavairoto īpatņu nekā sterilo kukaiņu tehnoloģijas (sterile insect technique) gadījumā, kas plaši tiek izmantota Vidusjūras augļmušas Ceratitis capitata ierobežošanai (Buchman et al 2018). Sterilo kukaiņu tehnoloģija ir videi draudzīga kukaiņu apkarošanas metode, kas ietver mērķa kaitēkļa masveida audzēšanu un sterilizāciju, izmantojot starojumu, kam seko sistemātiska sterilu tēviņu izplatīšana noteiktā vietā, kur tie pārojas ar savvaļas mātītēm, kurām šādas pārošanās rezultātā nav pēcnācēju, tādējādi kaitēkļu populācija samazinās (https://www.iaea.org/topics/sterile–insect–technique).
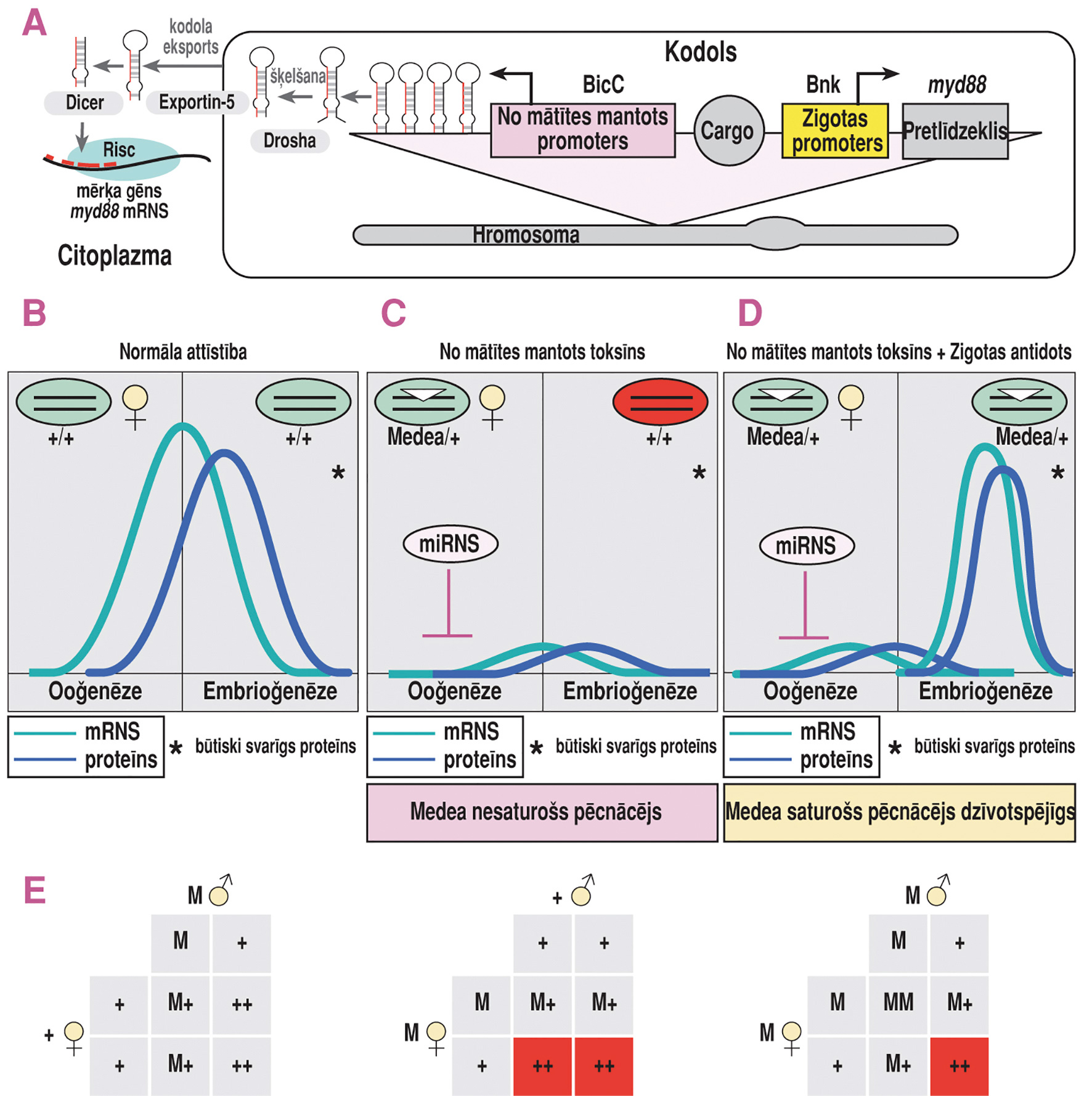
1. attēls. Mākslīgā Medea sistēma D. suzukii. (A) D. suzukii Medea transgēns tika izveidots, lai saturētu miRNS toksīnu, kas iedarbojas uz gēna myd88 5’ UTR sekvenci, kas ekspresiju nodrošina specifisks no mātītes pārmantots BicC promoters, un pretlīdzekli, kas sastāv no D. suzukii myd88 kodējoša reģiona, kura ekspresiju nodrošina agrīnai embrioloģiskajai stadijai atbilstošs Bnk promoters, kā arī divus atsevišķus transformācijas marķierus. Šie marķieri ir eGFP, kuru kontrolē acu krāsai specifisks 3xP3 promoters, un dsRed, kuru kontrolē bieži sastopams hr5–IE1 promoters. (B) Normālas attīstības gadījumā no mātītes mantotais myd88 tiek uzkrāts embrijā, kur tas ir nepieciešams normālai attīstībai. (C) Medea sistēmas miRNS toksīns iedarbojas uz myd88 mRNS ooģenēzes laikā, kavējot normālu uzkrāšanos embrijā un izraisot embrija nāvi tiem pēcnācējiem, kas nav pārmantojuši Medea sistēmu. (D) Embrijiem, kuriem ir Medea sistēmas kopija, ekspresējas tāds myd88 variants, kas nav jutīgs pret miRNS toksīnu, kas ekspresējas agrīnās embrioģenēzes stadijās, kas rezultējas ar miRNS izraisītu letalitāti. (E) Ja krusto heterozigotiskus Medea vīrišķos īpatņus ar savvaļas tipa sievišķajiem īpatņiem, visi pēcnācēji izdzīvo, jo no sievišķā īpatņa mantotais toksīns netiek ekspresēts (kreisajā pusē). Ja krusto heterozigotiskus Medea sievišķos īpatņus ar savvaļas tipa vīrišķajiem īpatņiem, 50 % pēcnācēju, kas nav pārmantojuši Medea sistēmu, iet bojā (vidū). Krustojot heterozigotiskus sievišķos īpatņus ar heterozigotiskiem vīrišķajiem īpatņiem, 75 % pēcnācēju pārmanto Medea sistēmu vai nu no tēva, vai mātes un izdzīvo, kamēr tie, kas nepārmanto, neizdzīvo (labajā pusē).
Attēls adaptēts no Buchman et al (2018). Paskaidrojumi attēlā redzamajiem apzīmējumiem: Dicer – endoribonukleāze Dicer, kas šķeļ pre–microRNS īsākos fragmentos, kurus sauc par microRNS (miRNS); Risc – RNS inducēts gēna izslēgšanas komplekss; Drosha – ribonukleāze, kas darbojas microRNS processinga sākuma stadijās. Exportin–5 – proteīns, kas eksportē pre–microRNS ārā no kodola uz citoplazmu; Cargo – jebkurš mērķa gēns, ko var ievietot Medea sistēmā.
Līdz ar Medea sistēmu eksistē vēl arī citas gēnu dziņu sistēmas, piemēram, tādas, kas balstītas un CRISPR/CAS sistēmu. Tā ir pašlaik plaši izmantota gēnu rediģēšanas sistēma, ar kuras palīdzību rada endonukleāžu dziņu sistēmu. Šīs sistēmas priekšrocība ir tāda, ka tiek izmantota viegli izveidojama vadības RNS (guide RNA), kas atrod genomā savu vietu. Pamatelementi šai sistēmai ir CRISPR endonukleāzes gēns, viena vai vairākas vadības RNS sekvences un atkarībā no pielietošanas veida mērķa gēns (2. attēls). Sistēmu parasti ievieto plazmīdā, kurā abās pusēs dziņa kasetei ir mērķa sekvencei homologas sekvences, lai inducētu integrēšanos genomā (homing) (COGEM, 2018). Ar šīs sistēmas palīdzību ir radītas tādas mutācijas, kas izraisa izmaiņas sievišķo īpatņu dzimumorgānos, tādējādi ietekmējot vairošanos (Fang and Scott, 2016). Citos gadījumos dabā varētu palaist sterilus vīrišķos īpatņus, kas pārotos ar savvaļas sievišķajiem īpatņiem, tā rezultātā samazinot vai pilnībā iznīcinot punktspārnu augļmušas populācijas (Kalajdzic and Schetelig, 2017). Pamatā gēnu dziņa modifikācijas konkrētajā populācijā neuzskata par paliekošām jeb permanentām. Uzskata, ka vienmēr izselekcionēsies rezistentas alēles. Tomēr tas varētu prasīt ilgu laiku, kas būtu pietiekams, lai pārtrauktu konkrētā kaitēkļa populācijas attīstību (COGEM, 2018).
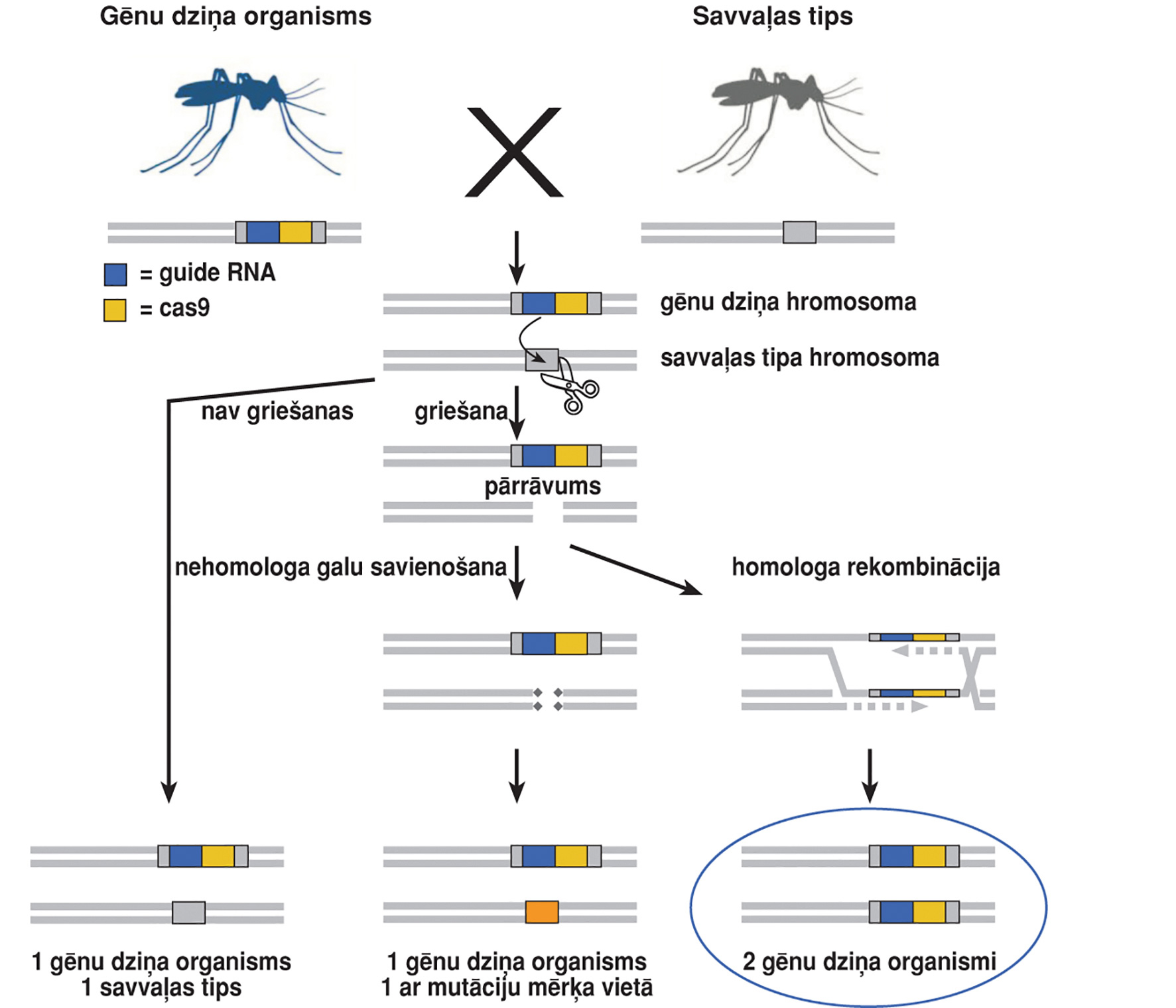
2. attēls. CRISPR/Cas gēnu dziņa izplatīšanās (attēls adaptēts no COGEM, 2018).
Šī jaunā tehnoloģija ir izraisījusi gan zinātnieku entuziasmu, gan dažādu ekspertu bažas. Kaut arī nākotnē gēnu dziņu sistēmas varētu izmantot lauksaimniecības kaitēkļu un invazīvu sugu kontrolei, apdraudētu sugu glābšanai vai slimību pārnēsātāju nomākšanai, tomēr vispirms nepieciešams novērtēt to iespējamās nevēlamās blakusparādības un izraisītās izmaiņas ekosistēmās. Eiropas pārtikas nekaitīguma iestāde pašlaik strādā pie gēnu dziņu organismu riska novērtējuma, iesaistot šajā procesā arī ES dalībvalstu iedzīvotājus, nevalstiskās organizācijas un kompetentās iestādes (EFSA, 2020).
Pateicības
Informācija sagatavota Lauku atbalsta dienesta finansētā projekta Ar jaunām ģenētisko modifikāciju metodēm iegūtu pārtikas, dzīvnieku barības un to piedevu noteikšana un šādu produktu zinātniskā riska novērtējums ietvaros.
Izmantotā literatūra





 Saimnieks LV
Saimnieks LVKS Piena Loģistika desmit gadu jubileju sagaida ar vērienīgu izaugsmi, no 2...
 Saimnieks LV
Saimnieks LVVeterinārārste Līga Spūle ikdienā strādā ar visdažādākajām lauku saimniecīb...
 Saimnieks LV
Saimnieks LVAudzējot eksporta apjomus, investējot ārpus sprostu vistu novietnēs vairāk ...
 Saimnieks LV
Saimnieks LVNogriežoties no Kandavas–Sabiles ceļa uz Matkuli, pēc nepilna kilometra ceļ...



